NEBridge® Golden Gate Assembly
Learn how to push the limits of your Golden Gate Assembly, and try one of our convenient kits (using BsmBI-v2 or BsaI-HFv2)
The efficient and seamless assembly of DNA fragments, commonly referred to as Golden Gate Assembly (1,2), has its origins in 1996, when for the first time it was shown that multiple inserts could be assembled into a vector backbone using only the sequential (3) or simultaneous (4) activities of a single Type IIS restriction enzyme and T4 DNA ligase. Golden Gate Assembly and its derivative methods exploit the ability of Type IIS restriction endonucleases (REases) to cleave DNA outside of the recognition sequence. The inserts and cloning vectors are designed to place the Type IIS recognition site distal to the cleavage site, such that the Type IIS REase can remove the recognition sequence from the assembly. The advantages of such an arrangement are three-fold:
- The overhang sequence created is not dictated by the REase, and therefore no scar sequence is introduced.
- The fragment-specific sequence of the overhangs allows orderly assembly of multiple fragments simultaneously.
- The restriction site is eliminated from the ligated product, so digestion and ligation can be carried out simultaneously.

The net result is the ordered and seamless assembly of DNA fragments in one reaction. The accuracy of the assembly is dependent on the length of the overhang sequences. Therefore, Type IIS REases that create 4-base overhangs (such as PaqCI®, BsaI-HF®v2, BbsI/BbsI-HF, BsmBI-v2 and Esp3I) are preferred. Insert assembly calls for careful design of overhangs to direct the assembly, as well as verification that the Type IIS REase sites used are not present in the fragments for the assembly of the expected product. The use of web tools such as the NEB NEBridge® Golden Gate Assembly Tool greatly simplifies both processes, making Golden Gate Assembly a robust technology that assembles single and multiple DNA fragments (5), even if repetitive elements are present (6) and can, if wished, introduce multiple site-directed mutations (7). Golden Gate Assembly has been widely used in the construction of custom-specific TALENs for in vivo gene editing (8), as well as in the cloning of inserts from diverse populations enabling library creation.
Recently, NEB has published research on T4 DNA Ligase Fidelity and multi-fragment assembly (9-12). This information, in conjunction with improved Type IIS restriction enzymes (e.g., BsaI-HFv2, NEB #R3733 and BsmBI-v2, NEB #R0739) and ligase fidelity tools, has enabled NEB to push the limits of Golden Gate Assembly. For more information, please visit www.neb.com/GoldenGate.
NEB has developed convenient kits (using BsmBI-v2 and BsaI-HFv2) for performing Golden Gate Assembly. For additional flexibility and convenience for users, NEBridge® Ligase Master Mix performs high efficiency and high-fidelity Golden Gate Assembly with a broad assortment of NEB Type IIS restriction enzymes.
Visit Type IIS Restriction Enzymes for a comprehensive list of all Type IIS enzymes available from NEB and their characteristics.
To help select the best DNA assembly method for your needs, please use our Synthetic Biology/DNA Assembly Selection Chart.

References:
- Engler, C., et al. (2008) PLoS ONE 3, e3647.
- Engler, C., et al. (2009) PLoS ONE 4, e5553.
- Lee, J.H., et al, (1996) Genet. Anal., e1647.
- Padgett, K.A. and Sorge, J.A. (1996) Gene, e7318.
- Werner, S., et al. (2012) Bioeng. Bugs, e18223.
- Scior, A., et al. (2011) BMC Biotechnol., e1187.
- Yan, P., et al. (2012) Anal. Biochem., e7029.
- Sanjana, N. E., et al. (2012) Nat. Protoc., e1431.
- Potapov, V. et. al. (2018) ACS Synth. Biol., e0333.
- Pryor, J. M. et al. (2020)PLoS One, e8592.
- Pryor, J.M., et al. (2022) ACS Synth. Biol. e0525.
- Sikkema A.P. et al. (2023) Current Protocols. e882.
Choose Product:
-

Golden Gate Assembly Workflow
-

Overview of NEBridge® Tools for Golden Gate Assembly
-

Function and Application of Type IIS Restriction Enzymes
-

NEB® TV Ep. 15 – Applications of Restriction Enzymes
-

Listen to DAD Informatics tools and NEB enzymes to enable complex one-pot Golden Gate Assemblies
-

Listen to DAD (Data-optimized Assembly Design) when constructing high-complexity Golden Gate Assembly targets
-

Restriction Enzymes in Golden Gate Assembly
-
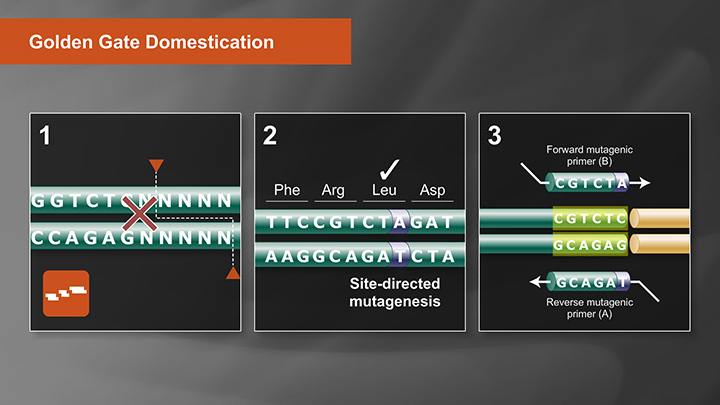
Golden Gate Assembly Domestication Tutorial
-

Golden Gate Assembly Tool Tutorial
