Gaussia and Cypridina Luciferases - Ultrasensitive secreted reporters and their use in dual assays
Scientists have long been interested in the biology of bioluminescent (i.e. light-emitting) organisms. Bioluminescence has evolved independently in a number of different phyla and is used by a wide range of marine and terrestrial organisms in behaviors as diverse as predator avoidance and sexual selection (1). Studies in this area led to the identification of multiple bioluminescent proteins and enzyme systems, some of which have found direct applications in biological research (2).
Luciferases are enzymes that produce light as a by-product of the oxidation of specific substrates called "luciferins". This light emission can be easily detected and measured, making luciferases ideal for the development of biological reporter systems. The advantages of luminescence, over fluorescence, include the absence of background, the amplification of signal and a high dynamic range that spans many orders of magnitude. Since light emission depends strictly on the chemical reaction between the substrate and the luciferase, there is no background light originating from the sample. Futhermore, the turnover of the light reaction significantly amplifies the reporter signal. Luciferases have become indispensable for the study of fundamental cellular processes including the regulation of transcription and translation, mRNA and protein stability, and nucleic acid/protein interactions. They also serve as markers of cellular physiology and responses (2).
The first generation of luciferase reporters developed were based on Firefly Luciferase (FLuc). Insect luciferases, including Photinus pyralis (firefly), catalyze the oxidation of a benzothiazol luciferin requiring ATP as a cofactor. Later, a marine luciferase from the sea pansy Renilla reniformis was isolated. Renilla Luciferase (RLuc), like most marine luciferases, uses a different luciferin, the benzylimidazo-pyrazinone coelenterazine. This feature allows measurement of RLuc activity from the same samples expressing FLuc.
Although both of these intracellular luciferases are widely used in life science research, they have the disadvantage of requiring lysis of the expressing cells in order to measure their activity. This becomes a disadvantage in many types of experiments. For example, measurement of activity at different time points involves experimental design with large numbers of wells and large amounts of reagents. Additionally, cell lysis precludes additional downstream procedures.
Recently, a new generation of bioluminescent reporters with unique characteristics that expand the flexibility and applications of luciferase reporter systems has become available. The secreted luciferases derived from Gaussia princeps, Metridia longa and Cypridina noctiluca do not require cell lysis to measure activity and are significantly "brighter" than firefly and Renilla. These features make these luciferases ideal for repeated assays, sensitive measurements in miniaturized assays and single cell applications.
Gaussia Luciferase
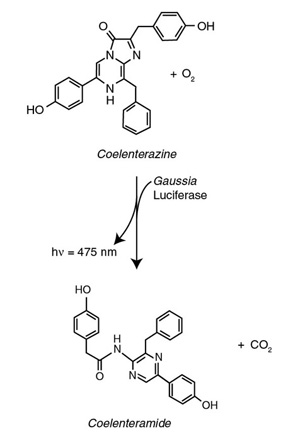
Naturally secreted from the deep sea copepod Gaussia princeps (3) (Figure 1), Gaussia Luciferase (GLuc) was first used in a sensitive analytical assay in 2002 (4) and later commercialized by NEB for expression in mammalian cells. GLuc is a monomeric enzyme of only 185 amino acids (19 kD), and is the smallest known luciferase. It catalyzes the oxidative decarboxylation of coelenterazine to produce coelenteramide via an excited state intermediate, which upon relaxation to the ground state emits blue light (475 nm) (Figure 2). The catalytic properties of GLuc make it an extremely sensitive detection reporter. Recombinant GLuc has been shown to produce the highest number of photons per mole of any luciferase (5). The first report describing the utility of GLuc in in vivo imaging demonstrated superior sensitivity over Renilla and Firefly by several orders of magnitude. Despite the fact that secretion causes its diffusion in the locality of the tissue that express it, the secreting cells were readily detectable due to this high sensitivity (6).
Figure 1: The copepod Gaussia princeps
Figure 2: The Photo-oxidation catalyzed by Gaussia Luciferase
The robust secretory signal, small size and sensitivity of GLuc, as compared to standard bioluminescent reporters, give GLuc some unique advantages in many assay systems. This is illustrated in the following examples.
The native secretion signal of GLuc has been shown to be functional in every eukaryotic system tested so far, from algae and nematodes to mice and mammalian cells (7,8). Functionality is dependent on the ER-Golgi pathway for proper folding and secretion. Thus the GLuc assay provides a fast, quantitative and sensitive technique to monitor the secretory pathway and ER stress (9,10).
The small size of Gaussia is favorable for achieving fusions with heterologous proteins without detrimental effects to either partner. Capul and de la Torre developed an elegant assay for viral infectivity using such a fusion. GLuc activity in the culture medium reflected the budding activity of the virus and the system was used for antiviral agent screening (11). In a different study, a chemokine- GLuc fusion was used to monitor interactions with its receptor by measuring cell-associated luciferase activity (12). Notably, the receptor was activated by the chemokine-GLuc chimera in a manner indistinguishable from the unfused chemokine. In both of these examples, the secretion signal was contributed by the fusion partner.
In research performed at NEB, scientists used Gaussia Luciferase activity secreted from cells subjected to RNAi, as a surrogate for cytoplasmic mRNA levels. The siRNA target sequence was introduced in the 3´-UTR of GLuc and the GLuc activity reflected the degree of silencing and the relative potency of the siRNAs tested (13). Despite the indirectness of the assay, reporter activity correlated well with RT-qPCR measurements of the mRNA levels, making the siRNA potency assay rapid, convenient and amenable to higher throughput formats.
Most reporter assays benefit by measuring a different reporter from the same experimental sample as a control. In the NEB study mentioned above, two additional reporters were measured from the cell lysates: beta-galactosidase and Firefly Luciferase. In a different study, GLuc was combined with Firefly Luciferase (FLuc) to study microRNA biogenesis (14). FLuc was used to monitor the activity of the miR-23 microRNA promoter, while GLuc was used to monitor the silencing activity of the mature microRNA. Although GLuc can also be measured in cell lysates, firefly, beta-galactosidase and Renilla activity can not be measured in the supernatant, making an additional secreted luciferase reporter desirable.
Cypridina Luciferase: a second secreted reporter
The first luciferase from Cypridinae ostracods was isolated from Vargula hilgendorfii and characterized several decades ago (15). This luciferase oxidizes a luciferin known as Vargulin, which is another common substrate of marine luciferases. Problems with substrate stability and the cost of production, however, precluded its wider application in analytical screening and cellbased applications. In 2004, a luciferase from a related species, Cypridina noctiluca, was reported to offer some advantages as a secreted reporter in mammalian culture medium (16). Furthermore, the development of synthetic approaches to produce Cypridina substrates resistant to rapid oxidation, helped establish this system as optimal for bioluminescent assays including reporter gene studies, live cell monitoring and BRET applications in vivo.
Cypridina Luciferase (CLuc) shares many of the features of Gaussia Luciferase, including secretion, emission of large amounts of light and high protein stability. Both GLuc and CLuc are inherently stable due to the presence of multiple disulfide bonds (4 and 17, respectively). This feature confers significant stability at high temperatures, making them useful for applications where other luciferases are rendered inactive. For example, both proteins maintain activity at 55ºC, a treatment that inactivates most viruses, or in the presence of 0.1 mM β-mercaptoethanol, a component of mouse embryonic stem cell culture media. In practice, this high stability allows the safe storage of luciferase containing extracts for later (re)measuring without any loss of activity.
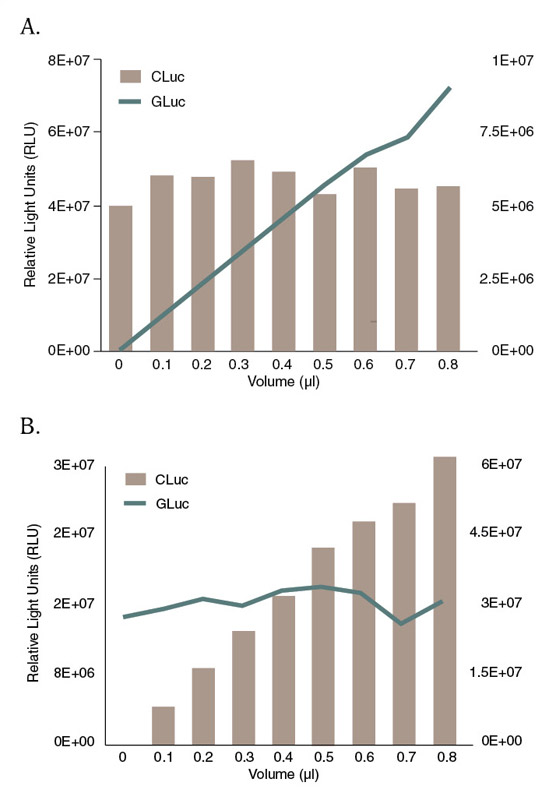
Perhaps the most compelling aspect of the CLuc reaction is its compatibility with GLuc in reporter assays, since GLuc and CLuc utilize two different substrates (Figure 2,3). The detection of one luciferase can be monitored without any cross-reactivity from the presence of the other in the same experimental sample (Figure 4). When expressed in mammalian cells, GLuc and CLuc are both secreted into the growth media, allowing the user to assay both luciferase activities from the same sample, multiple times over the course of an experiment. The use of these two secreted luciferase reporters in a dual secreted luciferase assay was first reported in 2007. This study demonstrated the activation of the promoter of the clock protein Bmal1, by the transcription factor RORa4 (17).
Figure 3: The photochemical reaction catalyzed by Cypridina Luciferase
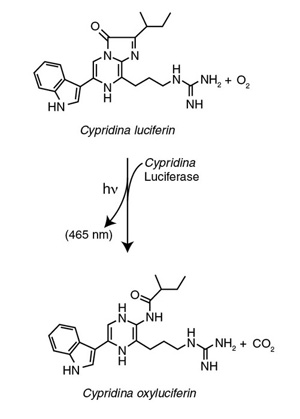
Figure 4: GLuc or CLuc activity can be measured without interference from the presence of the other luciferase
The ability to measure the activity of these ultrasensitive secreted luciferases from the same cells at different timepoints was featured in a study by Watanabe, et al (18). These investigators offered an excellent example of this property by showing that the cycling activity of clock genes could be monitored by continuously secreted GLuc and CLuc reporters over several days (18).
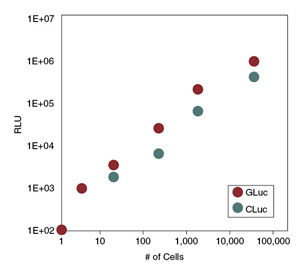
Figure 5: Detection of low numbers of cells expressing GLuc and CLuc
Summary
New England Biolabs now offers two compatible, secreted luciferases which can be used alone or together as a dual system. Gaussia Luciferase and Cypridina Luciferase are the brightest commercially available luciferases, enabling the user to detect activity in small samples or from very low expression (Figure 5). The non-destructive nature of these ultrasensitive assays allows the cells to remain intact so they can be used in further downstream assays including RT-PCR, Western blots, RNA expression analysis, live imaging, cell viability assays, etc. Their compatibility, secretion, sensitivity and stability, make GLuc and CLuc ideal for routine in vitro and in vivo reporter applications, as well as high throughput assays.
References
- Wilson, T. and Hastings, J.W. (1998) Annual Review of Cell and Developmental Biology, 14, 197–230.
- Roda, A., et al. (2009) Trends in Anal. Chem. 28, 307–322.
- Ballou, B., Szent-Gyorgyi, C. and Finley, G. (2000) Abstract from 11th Int. Symp. on Biolumin., & Chemilumin., Asilomar, CA.
- Verhaegen, M. and Christopoulos, K. (2002) Anal. Chem. 74, 4378–4385.
- Goerke, A.R. and Loening, A.M. (2007) J. Biotechnol. 128, 705–715.
- Tannous, B.A., et al. (2005) Mol. Ther. 11, 435–443.
- Ruecker, O., et al. (2008) Mol. Genet. Genomics, 280, 153–162.
- Knappskog, S., et al. (2007) J. Biotechnol. 128, 705–715.
- Badr, et al. (2007) PLoS ONE, 2, e571.
- Hewett, J.W., et al. (2007) Proc. Natl. Acad. Sci. USA, 104, 7271–7276.
- Capul, A. and de la Torre, J.C. (2008) Virology 382, 107–114.
- Luker, K., Gupta, M. and Luker, G. (2009). Biotechniques, 47, 625–632.
- Morlighem, J.E., Petit, C. and Tzertzinis, G. (2007) Biotechniques, 42, 599–606.
- Lee, J.Y., etal. (2008) Nucl. Med. 49, 285–294.
- Shimomura, O., Johnson, F.H. and Masugi, T. (1968) Science, 164, 1299–1300.
- Nakajima, Y., et al. (2004) Biosci. Biotechnol. Biochem. 68, 565–570.
- Wu, C., Suzuki-Ogoh, C. and Ohmiya, Y. (2007) BioTechniques, 42, 290–292.
- Watanabe, T., et al. (2010) Anal. Biochem. 402, 107–109.
By George Tzertzinis, Ph.D., Ana Egaña, Ph.D. and
Cathy Shea, Ph.D., New England Biolabs, Inc.

